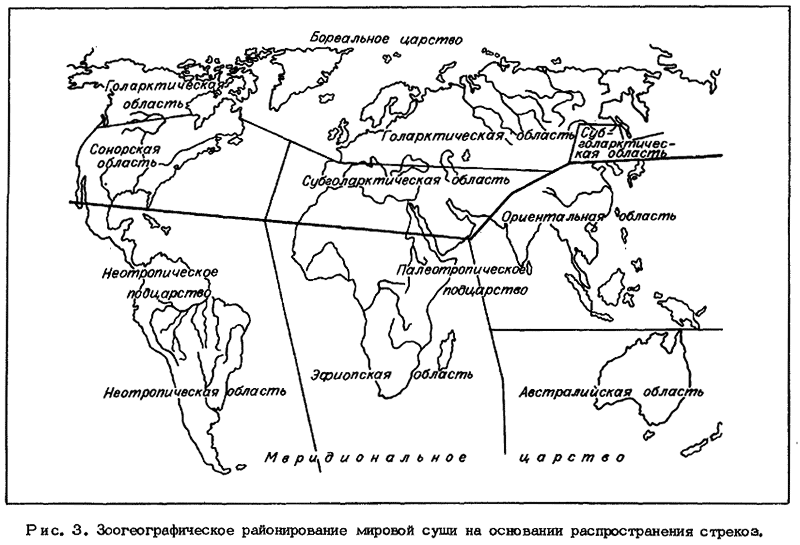
Фауна Бореального царства сложилась на территории Лавразии в относительной
изоляции от Гондваны, в пределах которой сформировалась фауна Меридионального
царства. При этом южная полоса Бореального царства - Сонорская область на
американском континенте и Субголарктическая на евразиатском - представлена
богатой и более древней одонатофауной, исходной для Северного полушария в целом.
Более обширная северная полоса Бореального царства - Голарктическая область -
отличается значительно более бедной и молодой по времени формирования фауной.
Особое значение в формировании бореальной фауны имеет выяснение возможных
миграционных путей, зависящих, в частности, от палеогеографической обстановки
соответствующих исторических периодов.
Однотипность голарктической фауны свидетельствует об очень тесных контактах и
исходном единстве американской и евразиатской одонатофаун. Сходство северной
биоты в целом и наличие у некоторых групп животных и растений так называемых
берингийских ареалов привели к признанию многими биогеографами Берингии как
моста для обмена фаунами между Северной Америкой и Азией и даже центра
формообразования голарктической фауны и флоры. Вероятно, для ряда групп, прежде
всего субарктических животных и растений, эти представления вполне обоснованны,
но далеко не для всех. Палеогеография Берингии во многом дискуссионна, хотя она
в определенные периоды представляла собой обширную сушу, климат в этом районе по
большинству реконструкций с конца палеогена был крайне суровым. Это явление
обусловлено глобальной дифференциацией климата в кайнозое и смешением
климатических зон, связанном с перемешением Северного полюса по приберенгийской
территории к его современному местонахождению.
Очевидно, Берингия послужила мостом-фильтром, пропускавшим одних животных и
задерживающим других, более теплолюбивых, в том числе стрекоз. Действительно, в
современном распространении стрекоз нет ни одного факта, подтверждающего их
берингийские трансконтинентальные миграции, но есть такие, которые
свидетельствуют в пользу североатлантических фаунистических связей двух
континентов по былой климатически благоприятной суше, называемой иногда Арктидой
и соединявшей Северную Америку, Гренландию, Шпицберген, Северные Европу и Азию в
единое целое. Множество палеогеографических и биогеографических данных
подтверждают важную роль этого канала трансконтинентальной связи фаун и флор.
Изоляция Тетисом и объединение Арктидой обусловливают основной и наиболее
древний тип формирования и принципиальное единство бореальной одонатофауны.
Дальнейшее ее формирование произошло на фоне дифференциации и колебаний климата,
антропогеновых оледенений и морских трансгрессий, перекрывавших крупные регионы
евразиатской суши и приводивших к дизъюнкциям ареалов.
В пределах Евразии наиболее древние дизъюнкции ареалов, прослеживающиеся в
современной фауне, возникли еще в палеогене. К ним, в частности, относятся
значительные долготные разобщения ареалов родов Pyrrhosoma и Boyeria. Виды
первого из них встречаются в Западной Европе (P. nymphula) и Южном Китае (P.
tinctipennis), второго - в Западной Европе (В. irene) и Японии (В. maclachlani
). Представители рода Boyeria (В. grafiana и В. vinosa) обитают и в Северной
Америке, где они распространены широко. Вероятно, разрыв ареалов этих родов
начался еще в палеогене под воздействием обширных морских трансгрессий и
развития аридности и повышения континентальности климата в результате поднятия
Центральной Азии.
Очень своеобразен дизъюнктивный ареал рода Epiophlebia, который относится к
процветавшему в мезозое реликтовому подотряду Anisozygoptera, представленному в
настоящее время единственным родом с двумя видами: Е. superstes, обитающим в
Японии, и Е. laidlawi, известным из Гималаев. Интересно, что оба вида этих живых
ископаемых сохранились в горах и отличаются исключительной реобионтностью,
обитая в холодных горных потоках с относительно стабильными гидрологическими и
биотическими условиями.
Наиболее большой разрыв видового ареала наблюдается у Coenagrion hylas -
сибирского вида, имеющего изолированный локальный ареал в Альпах. Очевидно, это
ледниковое разобщение и остаточный европейский участок ареала имеют реликтовый
характер. Вообще ледниковое время наложило глубокий отпечаток на формирование
рецептной евразиатской одонатофауны. Резкие похолодания и затопления в течение
этого времени уничтожили аборигенную фауну Западной Сибири. По мнению
большинства палеогеографов, на протяжении большей части плейстоцена на
юго-западе Западно-Сибирской равнины существовал обширный слабопроточный водный
бассейн, возникший вследствие ледниковой запруды рек бассейна Оби. Уровень
бассейна поднимался до 100 м и более, в результате чего происходил сток вод в
бассейны Тургая и Арала в виде широкого пролива, полностью изолировавшего
европейско-уральские и сибирские части ареалов стрекоз.
Восстановлению фауны в периоды регрессий водного бассейна и после его
исчезновения препятствовали климатические колебания. В частности, в позднее
постгляциальное время этот процесс был невозможен вследствие развития
ксеротермической климатической фазы, во время которой произошло остепнение
территории, резкое сокращение количества пресноводных водоемов, т.е. мест
обитания стрекоз, и сильная минерализация оставшихся. Так, по мере удаления от
горной части Урала к Васюганью, где в это время развились лесостепные
пространства, резко возрастала засоленность озер. Даже в отложениях глубоких,
тектонического происхождения предгорных озер появились диатомовые водоросли,
характерные для солоноватых вод /Благовещенский, 1943/. Еще сильнее процесс
осолонения развивался в более южных районах, что создавало неблагоприятные
экологические условия для существования стрекоз.
Восстановление одонатофауны на этой территории стало возможным сравнительно
недавно и происходило преимущественно за счет миграций с запада, о чем
свидетельствует современная структура ареалов и принадлежность экземпляров
западно-сибирских стрекоз к европейским подвидам. При этом у рада видов так и не
произошло полного восстановления ареалов, и на пространстве между Приуральем и
Приобьем они не обнаруживаются совсем или прерывисто распространены в основном
по долинам рек Тобол, Ишим и Иртыш.
Одонатофауна Восточной Европы испытала, возможно, не стопь катастрофическое, как
в Западной Сибири, но все-таки сильное влияние похолоданий и затоплений
ледникового времени, Еспи равнины неоднократно перекрывались ледниковыми водами,
то горные территории оказывались в более благоприятных условиях, и населявшая их
фауна могпа хотя бы частично сохраниться с доледникового времени. Об этом
свидетельствует большее по сравнению с прилегающими равнинами видовое
разнообразие и наличие реликтовых видов стрекоз на Алтае, Урале и в Карпатах.
Например, в горах Алтая и возвышенных приалтайских землях сохранились реликтовые
изолированные местонахождения видов Libellula depressa, Anax parthenope,
Sympetrum croceolum. В примыкающих к Алтаю ленточных борах бассейна Оби (также,
кстати, сохранившихся с неогена) сосредоточены популяции видов Leucorrhinia
pectoralis, L. albifrons, L. caudalis, Somatochlora flavomaculata и некоторых
других, основные ареалы которых расположены в европейской части континента.
Сходная ситуация наблюдается на Урале. Так, в предгорьях и горах Южного Урала
насчитывается 53 вида стрекоз в отпичие от 35 на сопредельных равнинных
территориях, причем уральские популяции ряда из этих видов изолированы от
основных ареалов.. К ним относятся, например, Brachytron pratense, Anax
parthenope, Somatochlora graeseri, Ischnura aralensis и некоторые другие.
Еще более заметные спеды ледниковых разобщений и последующей аридизации климата
обнаруживаются в широтных дизъюнкциях ареалов. Значительное распространение
среди евразиатских стрекоз имеют ареапы бореалъно- и даже арктоальпийского типа,
когда изолированные участки ареалов локализуются в южных горах, где сохраняются
популяции видов, широко распространенных на более северных широтах. Один из
наиболее ярких примеров такого разобщения - распространение стрекозы
Somatochlora sahlbergi. Основной ареал этого вида лежит в субарктической полосе
Евразии и Америки, а изолированные места нахождений известны в горах Южной
Сибири от Алтая до Прибайкалья.
В горах Таджикистана отмечено не менее 10 видов стрекоз, среднеазиатские участки
ареалов которых изолированы от основных, более северных областей
распространения. Примерно столько же южных горных иэолятов известно для Кавказа.
Большая часть этих видов в изолированных горных очагах морфологически не
отличается от популяций из основного ареала, но в отдельных случаях, как
например у Leucorrhinia dubia, они образовали особые подвиды.
Изредка наблюдается и обратное явление - изолированность на севере фрагментов
ареалов южных видов, представляющих собой, по- видимому, реликты
ксеротермических периодов. Иногда они связаны с горячими источниками, например
Orthetrum albistylum, основной ареал которого лежит в пределах Субголарктической
области, а северные изолированные очаги известны на выходах термальных вод на
северо-востоке Байкала и в долине р. Чары.
Относительно мало были затронуты ледниковыми пертурбациями в Бореальном
фаунистическом царстве территории Субголарктической области, что повлияло на
сохранение здесь богатой, разнообразной и достаточно древней одонатофауны. Так,
из 19 эндемичных для царства монотипических родов 18 оказались сосредоточеными в
Субголарктической области и лишь 1 - в Голарктической, крупнейшей по площади, но
наиболее молодой и бедной по видовому составу фаунистической области Земли.
Следует отметить, что все построенные на основании распространения стрекоз
принципиальные зоогеографические схемы и масштабные по времени
историко-фаунистические реконструкции хорошо согласуются с теорией
горизонтальных перемещений материков. Учитывая геологическую древность отряда
стрекоз, следует признать, что отражение в их распространении долговременных
геофизических процессов вполне естественно. Вместе с тем апелляции к идеям
мобилизма в зоогеографических работах вызывают иногда критические замечания.
Особенно энергично высказаны они в рецензии К.Ю. Еськова /1984/ на монографии по
географии стрекоз /Белышев, Харитонов, 1981, 1983а/. К сожалению, рецензия мало
касается сути монографий, явствующей уже из их названий, а именно географии
стрекоз, и сводится главным образом к критике палеогеографических основ
исторических реконструкций, занимающих в этом труде подчиненное положение и
составляющих незначительную часть его объема. Рецензент упрекает авторов в
приверженности к теории Вегенера, почему-то беря ее в кавычки, хотя здесь же
сообщает, "что ныне среди биогеографов предметом обсуждения является не факт
перемещения материков (признаваемый почти всеми), а масштабы и характер его
влияния на современное распространение живых организмов". От внимания
рецензента, видимо, ускользнуло то обстоятельство, что авторы подходят к данной
теории не как к непогрешимой догме и отдают себе отчет в том, что не все в ней
бесспорно, делая, например, такие замечания: "Конечно, на теорию А. Вегенера и
теперь приходится смотреть как на принципиальную схему, где необходима доработка
многих деталей, уточнение частностей и во времени, и в пространстве..."
/Белышев, Харитонов, 1981, с. 125/. "...Следует подчеркнуть и то обстоятельство,
что классическая схема А. Вегенера не отличается точностью. Она в принципе
верна, удостоверена данными геологии, палеомагнитологии, палеонтологии и
биогеографии, но, конечно, требует дальнейших доработок" /Там же, с. 128/. 64
Вместе с тем в литературе, в том числе и наиболее современной, существует столь
большой и общеизвестный разнобой в датировках и оценке масштабов движения земной
коры, что "все это заставляет нас отдавать пока предпочтение пиниям дрейфа Вегенера - Кеппена, построенным в основном на палеонтологическом материале" /Там
же, с. 125/. Казалось бы, авторская позиция оговорена достаточно четко, тем не
менее рецензент отмечает, что "авторы используют в конкретных построениях
картину перемещения материков и полюсов, воссозданную самим Вегенером, между тем
последние десятилетия внесли столь существенные коррективы в первоначальные
представления о континентальном дрейфе (см., например, /Красилов, 1977; Уэда,
1980/), что пользоваться реконструкциями 1924 г. по меньшей мере странно".
Кстати, В.А. Красилов, к работе которого апеллирует рецензент, отмечает, что
"теория Вегенера открыла новую страницу исторической биогеографии. Гипотеза
дрифта континентов органически вошла в биогеографические построения многих
исследователей... По современным представлениям, дрифт континентов - это частный
аспект тектоники плит" /Красилов, 1977, с. 63-64/.
Принципиальную правильность построений А. Вегенера даже на фоне самых
современных данных подчеркивают многие авторы. Так, Д. и М. Тарлинг /1973, с.
15/ отмечают, что "схема реконструкции Вегенера основывалась на фактах из многих
областей науки; она удивительно сходна с нашими современными представлениями об
эволюции южных материков". Известные советские геологи С.А. Ушаков и Н.А.
Ясаманов в капитальной сводке "Дрейф материков и климаты земли" /1984, с. 5/
подчеркивают, что «Теория тектоники литосферных плит представляет собой
современное развитие того направления науки о Земле, которое родилось еще в
начале XX в. в виде гипотезы дрейфа континентов. Факты, приводившиеся
мобилистами 50-60 пет назад в защиту "сумасбродной" гипотезы, выдержали проверку
временем; ныне эти факты получили полное подтверждение и дальнейшее развитие».
Академик А.. Л. Яншин в одном из своих выступлений в прессе отметил, что "сейчас
гипотеза Вегенера, получившая в ее современном виде название теории движения
тектонических плит, или новой глобальной тектоники, принята, с незначительными
поправками, геологами всего мира, лишь немногие ученые упорствуют в своем с ней
несогласии" /Известия, 08. 08. 84 г./.
Критикуя выводы авторов по генезису фауны, рецензент не обратил внимания на то,
что они даны как сугубо предположительные построения и во введении к книгам
подчеркнуто, что "в основу историко- фаунистического анализа положено
современное распространение стрекоз, морфологическая близость таксонов и
палеогеографические данные, которые опираются на теорию континентальных
перемещений, основы которой были заложены А. Вегенером... В вопросах истории и
времени расселения много условностей, поскольку палеонтологический материал
почти отсутствует и мы должны опираться на косвенные данные и логические
построения. Следовательно, все эти вопросы пока находятся на уровне рабочих
гипотез. Но высказать эти гипотезы мы должны, так как это продвинет изучение
вопроса независимо от того,
будут они приняты в свете новых данных или отвергнуты. То и другое равноценно,
так как подтолкнет к подобным исследованиям, даст для них опорные точки"
/Белышев, Харитонов, 1981, с. 4/.
В начале рецензии сообщено, что "мы посвящаем рецензию наиболее крупным
зоогеографическим обобщениям, выдвигаемым авторами", в ней в основном дан разбор
частных палеогеографических вопросов, причем далеко не во всем аргументация
рецензента бесспорна и точна. Например, рецензент пишет, что "авторы говорят о
существовании Пангеи еще в середине третичного периода... тогда как по
современным представлениям раскрытие Тетиса, отделившего Лавразию от Гондваны,
и одновременный раскол последней на восточную и западную части произошли уже в
конце триаса - начале юры". Такой упрек основан на явном недоразумении. В
разделе рецензируемой книги, посвященном общим вопросам формирования
голарктической фауны, написано: "История Северного полушария в свете теории А.
Вегенера рисуется следующим образом. До юры существовала общая континентальная
глыба. С этого времени она начала располагаться, ее части раздвигались, таким
образом наметился Атлантический океан" /Там же, с. 156/. Ясно сказано, что
целостность Пангеи сохранялась до юры, но никак до середины третичного периода,
применительно к которому речь идет уже лишь о фрагментах бывшей Пангеи.
Упрекая авторов в излишней приверженности к теории Вегенера в том, что они
"всецело полагаются" на его реконструкции, рецензент сокрушается по поводу того,
что в необходимых случаях они обращаются к современной литературе, уточняющей
схемы Вегенера. Так, К.Ю. Еськов пишет, что "для обоснования сильной
обособленности одонатофауны Индии и Юго-Западной Азии они пользуются
современными представлениями о северном дрифте Индостана как фрагмента Гондваны
и о его совсем недавнем контакте с Азией, что не согласуется с построениями
Вегенера, у которого Индия движется как часть Евразии".
Не вдаваясь более в дискуссию по достаточно частным и далеко выходящим за рамки
биологического труда проблемам, в заключение еще раз отметим, что все наши
зоогеографические и историко-фаунистические выводы базируются прежде всего не на
отвлеченных рассуждениях и не на спорных палеогеографических реконструкциях, а
на неоспоримых фактах распространения стрекоз, при этом они ни в коей мере не
претендуют на всеобщее зоогеографическое значение.
| © Стрекозы
Южного Урала, 2009-2010 © REDBOOK.RU, 2009-2010 |
|