Изучение суточных ритмов активности стрекоз - одно из необходимых условий для
оценки их роли в трофических сетях биоценозов, например, в регуляции численности
гнуса или вредителей сельского и лесного хозяйств, а также представляет интерес
с точки зрения выяснения адаптивных возможностей этих насекомых.
Крупные размеры и особенности поведения стрекоз делают их удобным модельным
объектом в полевых исследованиях суточных ритмов. Тем не менее, на общем фоне
публикаций по суточной активности насекомых стрекозам уделяется очень мало
внимания. Отрывочные сведения об изменении активности в течение дня получены для
нескольких европейских видов /Corbet, 1957; Parr, 1973/. В СССР исследования по
суточной активности стрекоз и влиянию на нее погодных условий проведены на о.
Кунашир /Заика, 1980/, в Южной Сибири /Белышев, 19676, 1974а; Заика, Воронова,
1977/, Восточном Приуралье /Харитонов, 1975а, б/, на юге Украины /Олигер, 1980,
1985/ и юго-западе Таджикистана,/Борисов, 1985в, 1987/. Накопленный материал
фрагментарен и невелик по объему, тем не менее позволяет обобщить имеющиеся
сведения и опыт в изучении суточной ритмики стрекоз, что может послужить
предпосылкой дальнейшего развития исследований в этом направлении.
В своих исследованиях мы придерживались терминологии, предложенной в монографии
B.Б. Чернышева /1984/ "Суточные ритмы активности насекомых", - первой сводке в
мировой литературе. Прежде чем перейти к обсуждению методов изучения суточной
активности, поясним, что она означает по отношению к стрекозам (рассматривается
только для имаго). По определению В .Б. Чернышева, активность- это любая
деятельность насекомого. Средством для ее осуществления служит подвижность. Б.Ф.
Белышев /19676/ признаком активности стрекоз считает полёт, так как с ним
целиком связано питание и размножение - важнейшие жизненные функции.
Представления о лёте и активности отождествляют и другие авторы /Заика,
Воронова, 1977; Заика, 1980; Олигер, 1980, 1985; Борисов, 1985в, 1987/, что в
целом неоправданно сужает диапазон понятия активности стрекоз, фактически
ограниченный периодом с момента начала движения после ночного покоя до полного
прекращения двигательной активности перед ночевкой. Иначе, между началом или
прекращением активности и непосредственно лётом существуют переходные периоды,
когда насекомые проявляют двигательную активность (например, выползают из
укрытий), но из-за действия каких-либо факторов еще или уже не способны к
полету. Эти периоды соответствуют времени потенциальной готовности - ВПГ
/Чернышев, 1973, 1981, 1984/. Знание ВПГ имеет непосредственно практическое
значение при профилактике гельминтозных заболеваний, переносчиками которых
являются стрекозы. Во время потенциальной готовности стрекозы наиболее уязвимы и
могут в массе поедаться птицами.
Суточный ритм складывается из разных видов частной активности. Например, летом
возле водоемов обычно наблюдается репродуктивная активность: охрана участков и
поиск или поджидание самок самцами, спаривание, откладывание яиц, выход имаго из
личинок. При этом у многих разнокрылых стрекоз, например видов рода Crocothemis,
Orthetrum, более активны и учитываются в основном самцы (до 98%), самки же
благодаря скрытному поведению и биотопической разобщенности с самцами в
отдельные часы суток отмечаются лишь единично. В удалении от водоемов
учитываемое соотношение полов близко к естественному и общее время активности
преимущественно трофической здесь больше, чем возле водоемов.
Некоторые виды частной активности наблюдать сложно, например, ночной выход имаго
из личинок, ночной охотничий лёт у Аnах parthenope Selys. Неясны пока причины
лёта на искусственный свет некоторых видов стрекоз. Таким образом, суточная
ритмика стрекоз — явление сложное, по всей вероятности, каждому виду активности
свойствен свой ритм и общая ритмическая картина складывается из их совокупности.
Несомненно при этом, что лёт стрекоз и связанные с ним виды активности имеют
основное значение в формировании суточных ритмов. Эндогенный суточный ритм
подвержен корректировке погодными факторами. Впервые количественная оценка
воздействия погодных факторов на активность стрекоз произведена Б.Ф. Белышевым
/19676/. А.Ю. Харитонов /1975а, б/ в дополнение к этому разработал методы
количественной оценки уровня активности.
В экспериментальных условиях с заранее заданными параметрами среды исследование
ритмики стрекоз сложно из-за их большой подвижности и проявлении эффекта
"замкнутого пространства" (повышенное возбуждение, стремление вырваться из
ограниченного пространства, затем потеря ориентации и, как следствие этого,
невозможность полёта). В полевых условиях необходимо параллельно учитывать
изменение уровня активности и изменение факторов среды.
Начало и прекращение активности можно фиксировать визуальными наблюдениями,
перемещаясь в местах ночевок или укрытий стрекоз при неблагоприятных погодных
условиях. Преимущество этого метода заключается в возможности наблюдать сразу
многие виды и на значительном пространстве. Основной недостаток - необходимость
отличать типичный лёт от спровоцированного наблюдателем во время потенциальной
готовности стрекоз. Более трудоемкий, но и более объективный метод - наблюдение
за поведением стрекоз на точно оконтуренных площадках или маршрутах.
На учетных площадках подсчитываются пролетающие стрекозы или количество
поведенческих актов в единицу времени (10-15 мин). Желательно отдельно учитывать
самцов, самок, копулирующие пары, самок, откладывающих яйца, и т.д. Наблюдения
за активностью мелких равнокрылых стрекоз целесообразно проводить на небольших
по размерам площадках (4-6 м), а учеты разнокрылых стрекоз - на площадках 6-10 и
более. Учеты на площадках более результативны в местах концентрации стрекоз, как
правило, возле водоемов. Показателем активности в этом случае служит количество
поведенческих актов (взлеты, патрульные облеты и др.), но последнее в сильной
мере зависит от количества активных особей, причем не только конспецифичных.
Известно, что с ростом плотности популяций до определенного предела у стрекоз
прогрессирует территориальное поведение /Мазохин-Поршняков, Рязанова, 1984;
Борисов, 1985в/.
На учетных маршрутах, протяженность которых определяется в зависимости от обилия
исследуемых видов (50-500 м), показателем уровня активности служит количество
активных особей в момент прохождения маршрута. Удобны кольцевые или параллельные
маршруты, проложенныее в местах, различающихся условиями, например, на солнце и
в тени, в заветрии и на открытом месте.
Изучение суточных ритмов невозможно без постоянного контроля за факторами среды.
Необходимо через определенные промежутки времени непосредственно в местах работы
учитывать время наблюдений и основные показатели: температуру и относительную
влажность воздуха, прямую солнечную радиацию, освещенность, давление, скорость
ветра, наличие осадков. Интервалы времени, через которые проводят наблюдения,
выбирают в зависимости от вариабельности показателей. Например, в моменты начала
и прекращения активности стрекоз и при резких изменениях погодных условий
наблюдения проводят с периодичностью 10-15 мин, в других случаях промежутки
можно увеличивать до 1 ч.
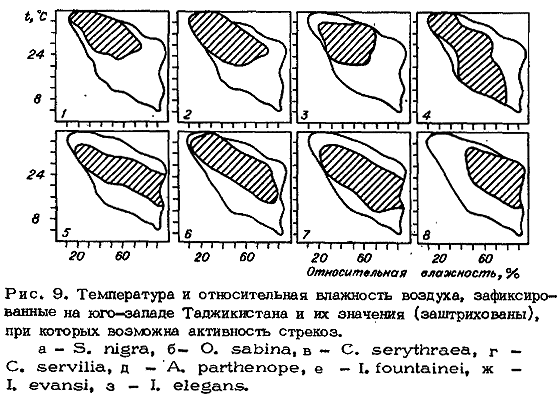 При обработке результатов наглядную картину дает графическое изображение
изменения показателя активности в сочетании с графиками изменения внешних
факторов. По оси абсцисс откладывается время суток, по оси ординат - показатели
уровня активности и факторов среды. При помощи таких графиков удобно проводить
анализ влияния факторов на ритм и уровень активности. Составление
термогигрограмм, совместных значений температуры и относительной влажности
воздуха, при которых зарегистрирована активность стрекоз, позволяет выделить
пороговые значения этих факторов и характеризует отношение к ним в общем
исследуемого объекта. Последнее возможно при достаточном количестве наблюдений
при условии большой вариабельности факторов. Например, при 600 наблюдениях на
юго-западе Таджикистана удалось построить такие термогигрограммы для восьми
видов (рис. 9).
При обработке результатов наглядную картину дает графическое изображение
изменения показателя активности в сочетании с графиками изменения внешних
факторов. По оси абсцисс откладывается время суток, по оси ординат - показатели
уровня активности и факторов среды. При помощи таких графиков удобно проводить
анализ влияния факторов на ритм и уровень активности. Составление
термогигрограмм, совместных значений температуры и относительной влажности
воздуха, при которых зарегистрирована активность стрекоз, позволяет выделить
пороговые значения этих факторов и характеризует отношение к ним в общем
исследуемого объекта. Последнее возможно при достаточном количестве наблюдений
при условии большой вариабельности факторов. Например, при 600 наблюдениях на
юго-западе Таджикистана удалось построить такие термогигрограммы для восьми
видов (рис. 9).
В настоящее время суточные ритмы стрекоз наиболее полно изучены в аридных
условиях на юго-западе Таджикистана. В зависимости от распределения максимумов
здесь выделяются три основных типа активности: преимущественно дневная - для
термоксерофильных видов на протяжении всего сезона и гигрофильных в холодное
время года (ранней весной, поздней осенью и зимой), преимущественно утренне-
вечерняя — для гигрофильных видов в теплое время года и преимущественно вечерняя
(сумеречная) - для ряда видов сем. Aeschnidae в весенне-летний период.
По отношению к гигротермическим факторам все виды стрекоз можно разделить здесь
на две группы: ксерофильную и гигрофильную. В группу ксерофильных видов входят
наиболее теплолюбивые виды стрекоз S. nigra, О. sabina, О. albistylum, О.
brunneum, С. erythraea, P. dealbata и эвритермный С. servilia. К этой группе,
вероятно, относится большинство разнокрылых стрекоз (исключая сем. Aeschnidae).
Максимум активности этих видов приходится на середину дня при наибольших
значениях температуры и низких влажностях воздуха (рис. 10, 1а).
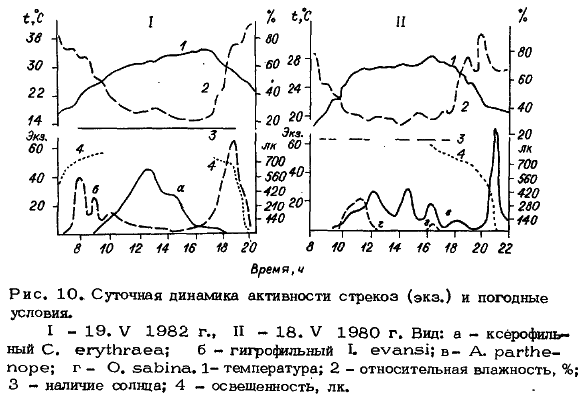 К гигрофильным видам относятся представители рода Ischnura (I. elegans, I.
evansi, I. fountainei ) и, видимо, еще некоторые равнокрылые стрекозы. Для видов
этой группы в теплый период года характерен бимодальный ритм с максимумами
активности в утренние и вечерние часы. Таким образом, они избегают воздействия
высоких температур при низкой влажности воздуха (рис. 10, 1б). К этой же группе
относится своеобразный по типу ритм Anax parthenope и, вероятно, некоторые
другие виды сем. Aeschnidae (Ае. coluberculus, Н. ephippiger) . у A. parthenope
наблюдаются два типа суточного лёта: репродуктивный и охотничий. Мощные вечерние
лики охотничьей активности приурочены к сумеркам и совпадают с максимумами
активности гнуса (рис. 10, II ). Отмечена также активность утром, в сумерках и
ночью, чего не наблюдается у стрекоз в умеренных широтах. Аналогичное поведение
у близкого вида A. imperator описывает Корбет/Corbet, 1957/ На юге Украины
вечерний охотничий лёт наблюдается у Brachytron. hafniense ( = pratense), Ae.
affinis, Ae. coluberculus, Cordulia aenea /Олигер, 1985/. Для видов рода Aeschna
активность с одним вечерним максимумом отмечена в лесостепи Восточного Приуралья
и Западной Сибири /Харитонов, 1975а, б/.
К гигрофильным видам относятся представители рода Ischnura (I. elegans, I.
evansi, I. fountainei ) и, видимо, еще некоторые равнокрылые стрекозы. Для видов
этой группы в теплый период года характерен бимодальный ритм с максимумами
активности в утренние и вечерние часы. Таким образом, они избегают воздействия
высоких температур при низкой влажности воздуха (рис. 10, 1б). К этой же группе
относится своеобразный по типу ритм Anax parthenope и, вероятно, некоторые
другие виды сем. Aeschnidae (Ае. coluberculus, Н. ephippiger) . у A. parthenope
наблюдаются два типа суточного лёта: репродуктивный и охотничий. Мощные вечерние
лики охотничьей активности приурочены к сумеркам и совпадают с максимумами
активности гнуса (рис. 10, II ). Отмечена также активность утром, в сумерках и
ночью, чего не наблюдается у стрекоз в умеренных широтах. Аналогичное поведение
у близкого вида A. imperator описывает Корбет/Corbet, 1957/ На юге Украины
вечерний охотничий лёт наблюдается у Brachytron. hafniense ( = pratense), Ae.
affinis, Ae. coluberculus, Cordulia aenea /Олигер, 1985/. Для видов рода Aeschna
активность с одним вечерним максимумом отмечена в лесостепи Восточного Приуралья
и Западной Сибири /Харитонов, 1975а, б/.
Строгого разграничения между различными по типу активности видами стрекоз нет. В
зависимости от конкретных погодных и микроклиматических условий ритмика у одного
и того же вида может закономерно изменяться. В холодное время года максимумы
активности всех видов стрекоз приходятся на самое теплое время суток.
В лесостепи Восточного Приуралья отмечено также три типа активности стрекоз
/Харитонов, 1975а, б/: с одним максимумом в середине дня (для большинства видов
при температуре не выше 28- 30 °С и влажности не ниже 30 %), с двумя пиками
активности в начале и конце дня (для большинства видов при температуре выше 30
°С и влажности воздуха менее 30 %) и с одним вечерним максимумом (для видов рода
Aeschna, охотящихся на скоплениях двукрылых насекомых). Следует отметить, что
принципиальных различий в суточных ритмах в пустынной зоне и лесостепи почти
нет, но на юге в жаркую середину дня активность некоторых видов падает почти до
нуля, а в лесостепи происходит ее незначительный спад.
В более северной таежной зоне стрекозам присущ лишь один тип активности с
максимумом в самое теплое время дня /Харитонов, 19756/. В лесотундре в условиях
непрерывной освещенности полярного дня и большой изменчивости других факторов
среды суточные ритмы стрекоз выражены слабо и описывающие их кривые отличаются
нетипичностью /Харитонов, 19756/.
На юге Украины А.И. Олигер /1980/ в зависимости от числа о обловленных
экземпляров на протяжении дня все виды стрекоз по типу активности распределяет
на пять групп: отловленные в основном в первой половине дня, более активные в
утренние и вечерние часы, с равномерной активностью на протяжении дня, с
максимумом активности в середине дня, с максимумом активности во второй половине
дня. Этот же автор /Олигер, 1985/ для ряда видов стрекоз описывает вечерние пики
активности. Метод отлова при изучении суточных ритмов по многим причинам
малопригоден применительно к стрекозам. Тем не менее по этим данным также
вырисовываются три типа активности, аналогичные таковым в лесостепи Восточного
Приуралья и пустынной зоне Таджикистана.
По наблюдениям В.В. Заики /1980/, на о. Кунашир выделяются виды с одним
максимумом активности в дневное время и с ее спадом в середине дня.
Таким образом, в южных районах СССР, где летние дневные температуры могут
превышать 28-30 °С, для стрекоз характерны три основных типа суточной
активности: с максимумом в середине дня, с двумя максимумами утром и вечером и с
одним вечерним.
Время активности стрекоз закономерно изменяется в зависимости от конкретных
условий среды, времени года и географического положения. На юго-западе
Таджикистана обшая продолжительность активности варьирует от практически
круглосуточной у A. parthenope в мае - июне до 5-7 ч у С. servilia и S.
decoloratum в декабре. У большинства термэксерофильных видов в теплый период
года максимальное время активности 15-16 ч в удалении от водоемов и 12-13
(репродуктивная активность) - возле водоемов, у гигрофильных 14-15 ч возле
водоемов. Период суточной активности у полисезонных видов (С. servilia, I.
elegans, Sympecma sp.) зимой примерно в 3 раза короче, чем летом.
В лесостепи Восточного Приуралья лёт большинства видов стрекоз длится около 15
ч, а у видов рода Aeschna превышает 17 ч /Харитонов, 19756/. В Южном Прибайкалье
при благоприятных погодных условиях лёт обычно продолжается не более 12 ч, а
максимальное время лёта не более 15 ч /Белышев, 19676/. По данным А.И. Олигер
/1980/, на юге Украины общая продолжительность лёта не превышает 14 ч, что
кажется несколько заниженным. В условиях островного климата на Дальнем Востоке
длительность лёта стрекоз значительно меньше, чем на материке, и составляет на
о. Кунашир 5-6 ч на побережье и 8-9 ч в кальдере вулкана, что обусловливается
высокой влажностью и сильными туманами в утренние и вечерние часы /Заика, 1980/.
Стрекозы в умеренных широтах - дневные насекомые. Исключение составляют
некоторые виды сем. Aeschnidae, активные также в сумерках и ночью. Из всех видов
двигательной активности полет больше всего нуждается в зрительной ориентации и
связан с определенными уровнями освещенности.
Освещенность — главный фактор, определяющий начало и прекращение активности,
имеет значение только при благоприятных гигро- термических условиях. Активность
большинства ксерофильных видов зарегистрирована при освещенности не менее
150-200 лк. Гигрофильные виды при прочих благоприятных условиях летают в
сумерках и активность их прекращается при минимальных значениях освещенности.
Сумеречная и даже ночная активность A. parthenope связана с активностью гнуса,
время роения которого четко связано с освещенностью /Чернышев, 1984/. Выход
имаго из личинок у многих видов происходит ночью, но первый полет - только
утром, при высоких ее уровнях.
В лесостепи Восточного Приуралья освещенность является сигналом к началу и
прекращению активности. Лёт стрекоз отмечен здесь при освещенности не менее
100-200 лк, а у видов рода Aeschnа - в глубоких сумерках при минимальных ее
уровнях /Харитонов, 1975а/.
В Южном Прибайкалье из-за неблагоприятных сочетаний других факторов в моменты
наблюдений лёт стрекоз отмечен только при высоких значениях освещенности
/Белышев, 19676/. Примерно также обстоит дело, судя по данным BJ3. Заики /1980/,
на о. Кунашир.
Температура и влажность воздуха - важнейшие факторы в регуляции суточных ритмов
стрекоз. Активность каждого вида ограничена определенными сочетаниями этих
показателей (см. рис. 9). Особенно сильно проявляется их воздействие на ритм в
пустынной зоне, где суточный размах температуры в местах обитания стрекоз
превышает 25 °С и значительно изменяется влажность воздуха (от 10 до 98 %).
Температурные пороги активности специфичны для каждого вида и значительно
отличаются. Так, например, активность С. servilia возможна в пределах от 7,2 до
39,6 °С (вероятно выше), A. parthenope- от 12,2 и только до 34 °С, S. nigra -
лишь от 23 до 40 °С и более, I. evansi - от 13 до 35,6 °С.
Температурные пороги лёта выше, чем пороги обшей подвижности, например у видов
рода Ischnura на 2-3 °С. Начало и прекращение лёта у гигрофильных видов
определяется преимущественно температурой, а у ксерофильных в значительной мере
и влажностью. Возле водоемов время лётной активности ксерофилов меньше на 2-4 ч,
чем у гигрофилов, и ограничено высокой влажностью в утренние и вечерние часы.
Изменение уровня активности на протяжении дня также находится в прямой
зависимости от гигротермических факторов. С увеличением температуры и падением
влажности (до известных пределов) снижается активность гигрофильных видов и
повышается у ксерофильных и наоборот. При чрезмерно высоких значениях
температуры (37-40 °С) и низкой влажности воздуха лёт гигрофилов прекращается, а
у самых теплолюбивых ксерофильных видов (например, S. nigra) уровень активности
максимален.
Температурные пороги активности у стрекоз на более северных широтах ниже, что и
естественно. Так, в Южном Прибайкалье лёт многих видов возможен при температуре
воздуха выше 10 °С /Белышев, 19676/, в лесостепи Восточного Приуралья - 13,5 °С
/Харитонов, 1975а/. Определяющее значение температуры (наряду с освещенностью) в
регуляции лётной активности отмечено в лесостепи и тайге, но особенно ярко
проявляется оно в лесотундре /Харитотов, 19756/. Высокая влажность воздуха и
тесно связанные с ней роса и туман, по данным Б.Ф. Белышева /19676/, оказывают
сильное сдерживающее влияние на активность стрекоз в Южном Прибайкалье. Такое же
явление отмечено для о. Кунашир /Заика, 1980/. В лесостепи Восточного Приуралья
при низкой влажности (<30 %) и высокой температуре ( > 28-30 °С) происходит
снижение уровня активности /Харитонов, 1975а/, аналогичное-таковому у
гигрофильных видов в пустынной зоне.
Прямая солнечная радиация. Особенно сильное влияние этого фактора на активность
стрекоз отмечено в пустынной зоне и в горах Средней Азии. В холодное время года
и при незначительном превышении температурой воздуха пороговых величин лёт
стрекоз возможен только при воздействии на них лучистой энергии солнца. Так,
зимой и ранней весной в пустынной зоне лёт С. servilia и S. decoloratum
возможен при температуре 7-1 О °С только в ясную погоду примерно с 11 ч при
достаточно высоком положении солнца. В пасмурные дни эти стрекозы не летают даже
при температуре воздуха 16-18 °С. В весенний период резко снижается уровень
активности ксерофильных видов в моменты, когда солнце скрывается за тучами (см.
рис. 10). В высокогорье Памира полностью прекращается лёт S. haritonovi, О.
brunneum и I. pumilio даже при незначительной облачности, не сопровождающейся
заметным падением температуры воздуха.
Летом в пустынной зоне при высокой температуре и низкой влажности воздуха прямая
солнечная радиация оказывает сдерживающее влияние на активность стрекоз. При
этом гигрофильные виды или полностью прекращают летать, или перемещаются в
затененные места. На более северных широтах отмечено, что облучение стрекоз
солнцем всегда содействует их активности и часто играет роль решающего фактора
начала лёта /Харитонов, 1975а/.
Воздействие ветра на активность стрекоз тесно связано с температурой воздуха.
Чем больше скорость ветра, тем большие значения температуры (и освещенности)
требуются для начала лёта. В лесотундре скорость ветра - один из главнейших
факторов в регуляции лётной активности /Харитонов, 19756/.
При высоких температурах воздуха сдерживающее активность стрекоз влияние ветра
значительно ослабляется. Так, в лесостепи стрекозы способны летать при скорости
ветра до 4,5 м/с /Харитонов, 1975а/, а в аридных областях Таджикистана отдельные
виды ( S. nigra) - да 8 м/с.
Отрицательное действие на активность стрекоз изменения атмосферного давления
отмечено в Южном Прибайкалье /Белышев, 19676/ и в лесостепи Зауралья /Харитонов,
1975а/. Даже в разгар лёта резкое падение давления (например, в течение 45 мин с
759 до 756 мм рт. ст) способно вызвать полное его прекращение.
В дождь обычно лёт стрекоз прекращается, но при слабых и кратковременных осадках
многие виды сохраняют активность.
В целом сведения о суточной активности стрекоз очень фрагментарны. Необходимо
изучение ритмики отдельных видов с учетом всех видов частной активности и
факторов, влияющих на них. Важно выяснить особенности ритмов стрекоз в
экстремальных условиях среды, например, в высоких широтах, в высокогорье, зимой
на юге, а у распространенна видов - в разных ландшафтно-географических районах.
В последнем случае, в частности, интересно выявить различия в формировании
суточных ритмов в разных частях ареалов у бореоальпийских видов.
Фауна
и экология стрекоз / Белышев Б.Ф., Харитонов А.Ю., Борисов С.Н. и др. -
Новосибирск: Наука. Сиб. отд-ние, 1989. - 207 с.
|